Article Text

Abstract
The Epstein-Barr virus (EBV) is detected in the tissue of about 10% of gastric carcinoma cases throughout the world. In each case, 100% of carcinoma cells are infected with EBV. Analysis of EBV in carcinoma biopsies indicates that carcinoma is formed by the proliferation of a single EBV infected cell. These findings suggest that EBV plays an important role in the development of EBV positive gastric carcinomas. The EBV genes expressed are EBV determined nuclear antigen 1 (EBNA1), two small non-polyadenylated RNAs known as EBER1 and EBER2, and the transcripts from the BamHI-A region (BARF0); in addition, some cases also express a small amount of latent membrane protein 2A (LMP2A). Epithelial cells are refractory to EBV infection in vitro. This has hampered the study of the role of EBV in epithelial malignancies. The use of recombinant EBV carrying a selectable marker has enabled this difficulty to be overcome. EBV infected cell clones can be obtained from most carcinoma cell lines examined, and it was found that cell to cell contact was an efficient mode of EBV infection. Furthermore, it was possible to immortalise primary gastric epithelial cells by EBV infection. The cells expressed identical EBV genes to those typically seen in EBV positive gastric carcinoma, and showed accelerated malignant properties, including growth in soft agarose and tumorigenicity in severe combined immunodeficient (SCID) mice. These results suggest that EBV contributes to the maintenance of the malignant phenotype of EBV positive gastric carcinoma.
- Epstein-Barr virus
- gastric carcinoma
- epithelial cell
Statistics from Altmetric.com
The Epstein-Barr virus (EBV), originally isolated from Burkitt's lymphoma biopsies, has now been linked to various epithelioid diseases.1 Hairy leukoplakia is a lesion caused by EBV replication. The very strong association between EBV and nasopharyngeal carcinoma is already well established (see G Niedobitek, this issue). Furthermore, the development of polymerase chain reaction (PCR) and in situ hybridisation (ISH) techniques revealed the association of EBV with many other malignancies, including gastric adenocarcinoma. About 10% of gastric carcinomas throughout the world are monoclonal proliferations of EBV infected carcinoma cells. Gastric carcinoma is one of the most common carcinomas, and the worldwide occurrence of EBV positive gastric carcinoma is estimated at more than 50 000 cases/year.
Unlike B cells, epithelial cells have displayed a remarkable resistance to EBV infection in vitro. This has hampered the study of the role of EBV in the development of epithelial malignancies. Therefore, we have established a system for the infection of epithelial cells in vitro, which has allowed us to study the role of EBV in the development of epithelial malignancies. Here, I review the literature concerning the association of EBV and gastric carcinoma, and introduce our recent findings obtained using our system for the infection of epithelial cells.
Epidemiology
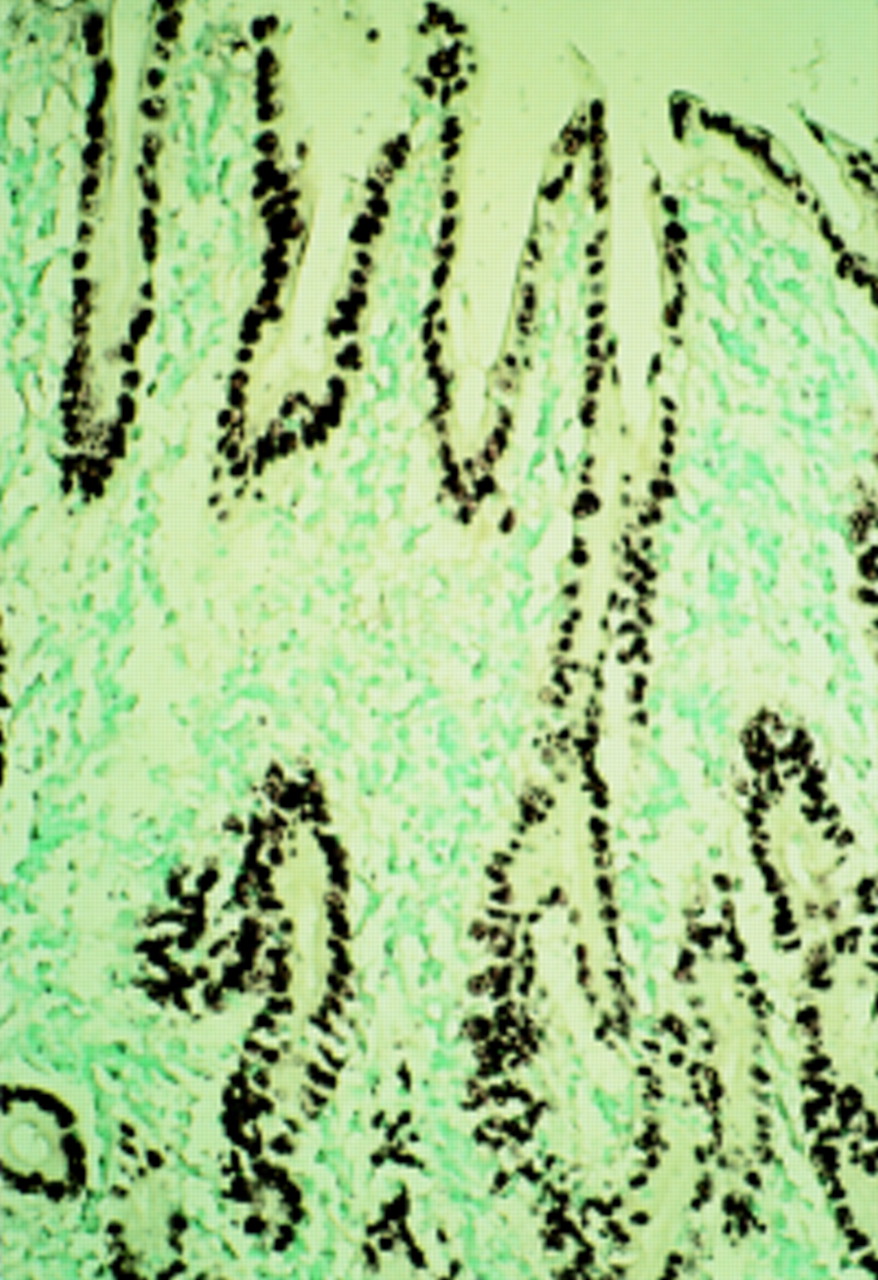
Most nasopharyngeal carcinomas are undifferentiated and accompanied by intense lymphoid infiltration (termed lymphoepithelioma). Carcinomas with a similar histological profile occur at a low incidence in organs such as the salivary glands, thymus, lungs, etc, mainly in Chinese and Inuits. These carcinomas are termed lymphoepithelioma-like carcinomas or carcinomas with lymphoid stroma, and most cases are EBV positive.1 The association between EBV and gastric carcinoma was first reported in this particular type of gastric carcinoma.2 EBV DNA was demonstrated in more than 80% of gastric carcinomas of the lymphoepithelioma type by PCR and ISH.3–6 Subsequently, Shibata and Weiss demonstrated EBV infection in gastric adenocarcinomas of ordinary histology (fig 1).7 They reported that EBV is present in almost all carcinoma cells in EBV positive cases. Interest has increased with a report that 69 of 999 (6.9%) cases of gastric carcinoma from Japan were EBV positive using ISH for the EBV encoded small RNAs (EBERs).8 Among them, the lymphoepithelioma type constituted only eight cases and the remaining cases were typical adenocarcinomas.

Detection of Epstein-Barr virus (EBV) encoded small RNA 1 (EBER1) in gastric carcinoma of the differentiated type. All carcinoma cells are positive for EBER1.
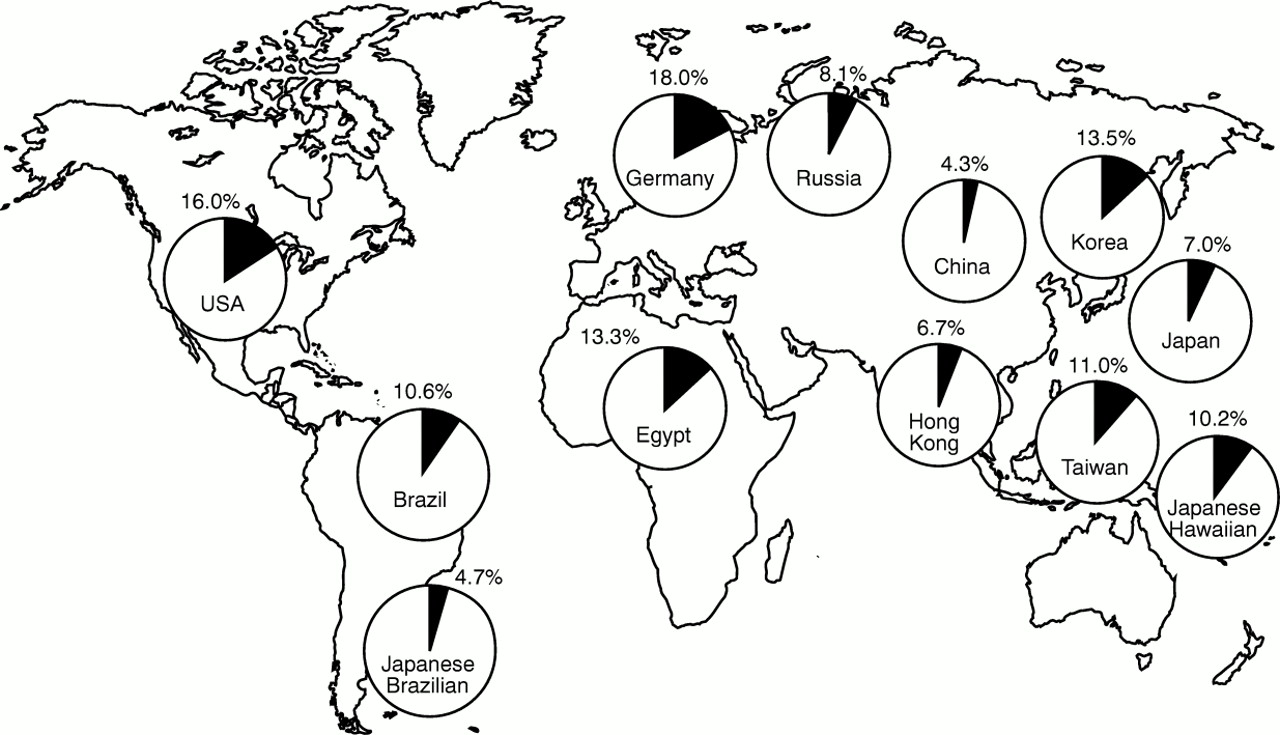
In contrast to Burkitt's lymphoma and nasopharyngeal carcinoma, which are endemic in equatorial Africa and Southeast Asia, respectively,1 EBV positive gastric carcinoma is a non-endemic disease distributed throughout the world. However, there are some regional differences in the incidence of EBV positive gastric carcinoma as a proportion of all cases of gastric carcinoma, with the highest (16–18%) in the USA and Germany, and the lowest (4.3%) in China (fig 2).7–16 Tokunaga et al examined the incidence of EBV positive cases in gastric carcinomas of nine Japanese cities.17 The highest incidence (10.3%) was observed in Okinawa, which has the lowest gastric carcinoma mortality rate in Japan (15.8/105 population), and the lowest incidence (3.1%) was in Niigata, which has the highest gastric carcinoma mortality rate (57.2/105). These observations suggest an inverse correlation between the incidence of gastric carcinoma and mortality from the disease. In Japan, the incidence of EBV positive gastric cancer is three times higher in men than in women, and is higher in younger men.8 EBV positive cases are more frequent in the cardia (8.1%), middle stomach (10.6%), and gastric stump (12.5%) than in the antrum (3.9%). EBV involvement is seen in papillary as well as tubular adenocarcinomas. There is no difference in EBV prevalence between intestinal and diffuse types of carcinoma.

World distribution of Epstein-Barr virus (EBV) positive gastric carcinoma. 16
Presence and expression of EBV in carcinoma cells and non-neoplastic mucosa
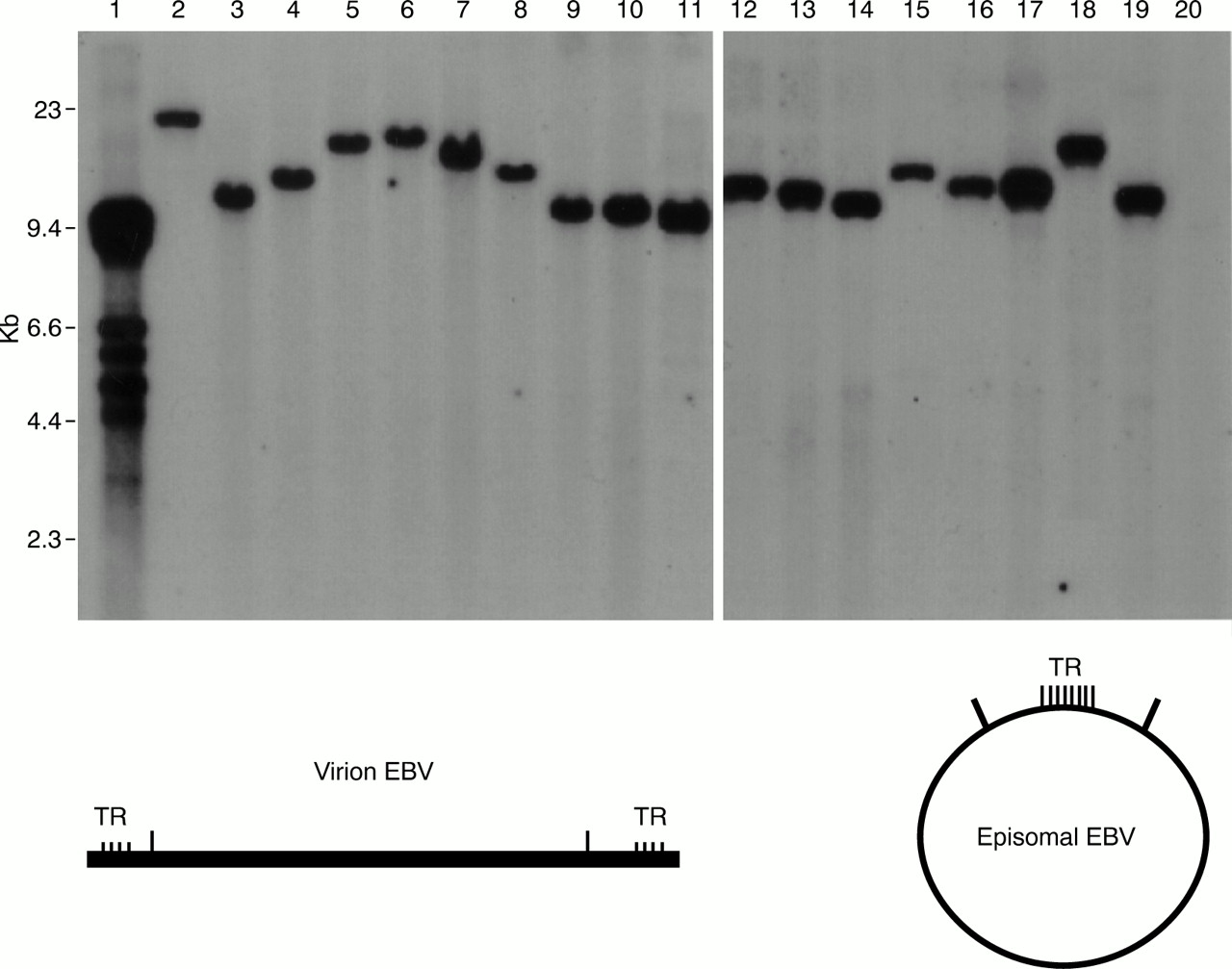
EBV is not integrated into the cellular DNA of carcinoma cells, but is maintained as a plasmid. The numbers of terminal repeats (TRs) in EBV positive gastric carcinomas are always uniform, and therefore the tumour is derived from a single EBV infected cell (fig 3).7,11,18,19 Taken together with the fact that 100% of carcinoma cells in EBV positive gastric carcinomas are EBV positive, this suggests that EBV plays an important role in the development of cancer. Of the six types of EBV determined nuclear antigens (EBNAs), only EBNA1 is expressed, and of the three latent membrane proteins (LMPs), LMP1 and LMP2B are not expressed, although LMP2A is expressed in some cases. In addition, the BARF0 gene from the BamHI-A region and the EBERs are always expressed.18,20 This EBV gene expression pattern is similar to that of Burkitt's lymphoma.

Epstein-Barr virus (EBV) DNA detection in EBV positive gastric carcinoma biopsies by Southern blotting. The EBV DNA terminal repeat (TR) was used as a probe. A single band was detected for each gastric carcinoma tissue, indicating that the gastric carcinoma was formed by proliferation of one EBV infected cell. Lane 1, EBV producing cell line B95-8 (multiple bands because the number of viral TRs is different for each virus particle); lane 2, Burkitt's lymphoma cell line Raji (single band); lanes 3–19, EBV positive gastric carcinoma samples (single bands); lane 20, EBV negative gastric carcinoma. Kindly provided by Dr S Imai, Kochi Medical School, Kochi, Japan.
In non-neoplastic gastric mucosa, scattered EBV positive cells are seen in dysplastic mucosa bordering the tumours by means of EBER ISH, but are absent in surrounding lymphocytes, other normal stromal cells, intestinal metaplasia, and normal gastric mucosa.7,9,12,14,19,21 These observations suggest that EBV infection occurs in the dysplastic phase and that an apparent growth advantage is conferred by the EBV infection.
Furthermore, a sensitive DNA ISH method revealed that non-neoplastic gastric epithelium, including intestinal metaplasia, is frequently infected with EBV, although EBER expression was not seen.22 A similar EBER negative EBV latency was found recently in the tissue of hepatocellular carcinoma.23 These observations highlight the danger of relying on EBER detection as the sole marker of latent EBV infection, and suggest that gastric epithelium might be a reservoir for EBV.
EBV specific immunity in patients with carcinoma
Patients with EBV positive gastric carcinoma have high IgG antibody titres against EBV capsid antigens (VCAs) and early antigens (EAs). IgA antibody against VCAs is detected in about 60% of cases,18,19 but its diagnostic value is limited because the titres are much lower than those in nasopharyngeal carcinoma. There is evidence of high antiviral titres before the diagnosis of EBV positive gastric carcinoma.24 These findings are similar to other EBV associated malignancies and suggest that active EBV infection exists before the development of EBV positive gastric carcinoma, although EBV specific cellular immunity is not greatly reduced.18
EBV associated gastric adenocarcinomas often accompany relatively strong lymphocyte infiltration. These tumour infiltrating lymphocytes (TILs) are predominantly human leucocyte antigen (HLA) class I restricted CD8 positive cytotoxic T lymphocytes (CTLs),25 which kill autologous EBV immortalised cells (but not phytohaemagglutinin blast cells).26 However, they do not recognise known EBV antigenic peptides presented by HLA molecules using fibroblasts infected with vaccinia recombinant virus expressing each of the EBV latent proteins.26 These data suggest that some EBV induced proteins might be involved in the strong T cell response, but that this response is not an effective way of killing EBV infected carcinoma cells.
Factors affecting carcinoma development
p53 mutation and overexpression are common in gastric carcinoma, and are detected not only in cancerous regions but also in areas of precancerous dysplasia and metaplasia.27 This suggests that p53 mutation might be an early event in gastric carcinogenesis. EBV positive gastric carcinomas tend to express much higher amounts of p53 compared with EBV negative cases.28 Leung et al have reported that nearly all EBV positive gastric carcinomas show weak to moderate p53 expression in a variable proportion of carcinoma cells, and suggest a role for EBV in a non-mutational mechanism of p53 overexpression.29 Such a hypothesis has also been proposed in nasopharyngeal carcinoma, in which p53 overexpression is detected in most cases yet sequence analysis has been unable to detect p53 mutation.30,31
The number of apoptotic tumour cells in EBV positive gastric carcinoma is lower than that in EBV negative cases.32,33 High bcl-2 expression in EBV positive gastric carcinoma might protect tumour cells from apoptosis.33
Helicobacter pylori infection induces chronic atrophic gastritis and subsequent intestinal metaplasia, and is linked epidemiologically with gastric carcinoma.34 Gastric carcinoma tissues are surrounded by atrophic mucosa and intestinal metaplasia in both EBV positive and EBV negative cases.35 It is conceivable that EBV infection occurs in the atrophic epithelial cells and leads to carcinoma development. There is no significant difference in the frequency of H pylori infection between EBV positive and EBV negative gastric carcinomas.35
After partial gastrectomy for benign diseases, carcinomas occur at a high frequency in the remaining stomach.36 This is referred to as gastric remnant cancer or stump cancer. The cancer risk increases with increasing length of the postoperative interval. Several explanations have been proposed for the high risk. Prolonged exposure to reflux of alkaline bile and pancreatic juices stimulates mucosal cell proliferation.37 The prevalence of EBV involvement in remnant carcinomas is significantly higher (27%) than in non-remnant carcinomas.38
The 30 bp deletion in the C-terminus of the LMP1 gene has been implicated in the pathogenesis of nasopharyngeal carcinoma and Hodgkin's disease. However, this deletion might not be relevant in the pathogenesis of EBV associated gastric carcinoma because high prevalence of the deletion in gastric carcinomas reflects the prevalence of the deletion variant in the normal population.39 The situation is similar for the EBNA1 variant with a deletion in the C-terminus.40
No specific linkage to HLA subtype has been reported in patients with EBV positive gastric carcinoma.41 EBNA3B is a major antigenic target that elicits an HLA-A11 CTL response to EBV infection. Mutations in the HLA-11 epitopes of EBNA3B are common in EBV associated gastric carcinomas, but are not related to the patient's HLA status.42
In vivo and in vitro models of EBV infection of epithelial cells
In addition to nasopharyngeal carcinoma and gastric carcinoma, many examples of EBV infection of epithelial cells in vivo are known, including AIDS related hairy leukoplakia, which is a proliferative EBV lesion in the lingual epithelium,43 and EBV infection of the salivary gland epithelium.44 However, in vitro, in comparison with the infection of B cells, infection of epithelial cells is more difficult. EBV adheres to B cells through CD21 molecules expressed on these cells, and this determines the B cell tropism.45 In contrast, epithelial cells are CD21 negative and this is believed to be why infection is not established, especially because they become susceptible to EBV infection when the barrier is overcome by CD21 expression via gene transfer46 or membrane implantation.47
We have shown clear evidence for direct infection of various human epithelial cells by EBV in vitro.48,49 The infection was achieved by the use of recombinant EBV (Akata strain) carrying a selectable marker gene,50,51 but without any other artificial operations, such as introduction of the CD21 gene. The cells used were 21 human carcinoma cell lines including those of the stomach, lung, and colon, normal human fibroblasts, and five non-human epithelial and fibroblast cell lines. Infection was accomplished by two methods: direct contact with high titre virus supernatant; and mixed culture with recombinant EBV producing Akata cells. After infection, cells were cultured in G418 containing medium and EBV infected cells were selected. Continuous EBV infected cell clones were obtained from three human carcinoma lines by the virus supernatant, and from 15 human carcinoma lines by the mixed culture method (table 1). In all of the continuously infected cell clones, only EBER, EBNA1, LMP2A, and BARF0 were expressed constitutionally .
Efficiency of Epstein-Barr virus (EBV) infection of epithelial cell lines
Sixbey and Yao have reported that in the pathway whereby IgA in the blood is processed into the secreted form by epithelial cells, EBV is incorporated into the epithelial cells in a bound state with IgA antibody against envelope protein.52 This phenomenon could explain the involvement of EBV infection in the development of nasopharyngeal carcinoma and possibly of gastric carcinoma, which are typically preceded and accompanied by the appearance of virus specific IgA in serum. Our data, however, point to another conceivable method of infection in vivo, in which cell to cell contact with virus producers is another efficient mode of EBV infection. The virus donors are most likely EBV infected B cells migrating into the epithelial stroma or intraepithelial space.
In our studies, most of the epithelial cell lines that were successfully infected with EBV were CD21 negative, and the infection was not inhibited by anti-CD21 monoclonal antibodies (OKB7). The results indicate that EBV infects epithelial cells through a receptor other than CD21. On the other hand, Fingeroth et al have reported that EBV can infect a human epithelial cell line, 293, in a CD21 dependent manner.53 These results suggest that EBV might be able to infect epithelial cells by more than one mechanism.
As in nasopharyngeal carcinoma, attempts to establish EBV positive gastric carcinoma cell lines have been unsuccessful. However, an EBV positive gastric carcinoma cell line that is transplantable in severe combined immunodeficient mice (SCID) mice (KT) has been established.54 The cell line retains the same clonal EBV genome and pattern of EBV gene expression as the original tumour biopsy. Thus, only the EBERs and Qp driven EBNA1 are expressed. This KT cell line could be an excellent in vivo model for studying the role of EBV in the development of gastric carcinoma. Two EBV positive epithelial cell lines have been established from non-cancerous portions of the stomachs of two patients with gastric carcinoma.55,56 Both cell lines express EBNA2 and LMP1, and spontaneously produce infectious viruses. These viruses are unique in that they have the ability to transform primary B cells and induce EA in latently EBV infected Raji cells. Apart from the P3HR-1 cell line, EA inducing virus has been reported in the nasopharyngeal carcinoma–KT cell line57 and a nasopharyngeal carcinoma cell line.58 The former is a nasopharyngeal carcinoma hybrid cell line obtained by fusing primary nasopharyngeal carcinoma cells with an epithelial cell line derived from a human adenoid tissue. EA inducing virus might therefore be more prevalent in epithelial tissues.
With regard to the virus strain dependent difference in infection efficiency suggested previously,46,59 our preliminary results indicate that the B95-8 strain of EBV is also infectious to epithelial cells. However, we have not determined the relative infection efficiencies of Akata and B95-8 viruses.
Growth promoting effects of EBV in epithelial cells
EBV immortalises B cells in vitro. EBNA2 and LMP1 appear to play the most important roles in the immortalisation of lymphocytes.60 However, these molecules are not expressed in EBV associated gastric carcinoma, raising doubts about the importance of the presence of EBV. We and others have analysed the Akata Burkitt's lymphoma cell line, which retains type I EBV latency in vitro, and have demonstrated that the presence of EBV is essential for the maintenance of the malignant phenotype and apoptosis resistance.61–64 Therefore, it is also possible that EBV contributes to the development and maintenance of gastric carcinomas, as described for Akata Burkitt's lymphoma cells.
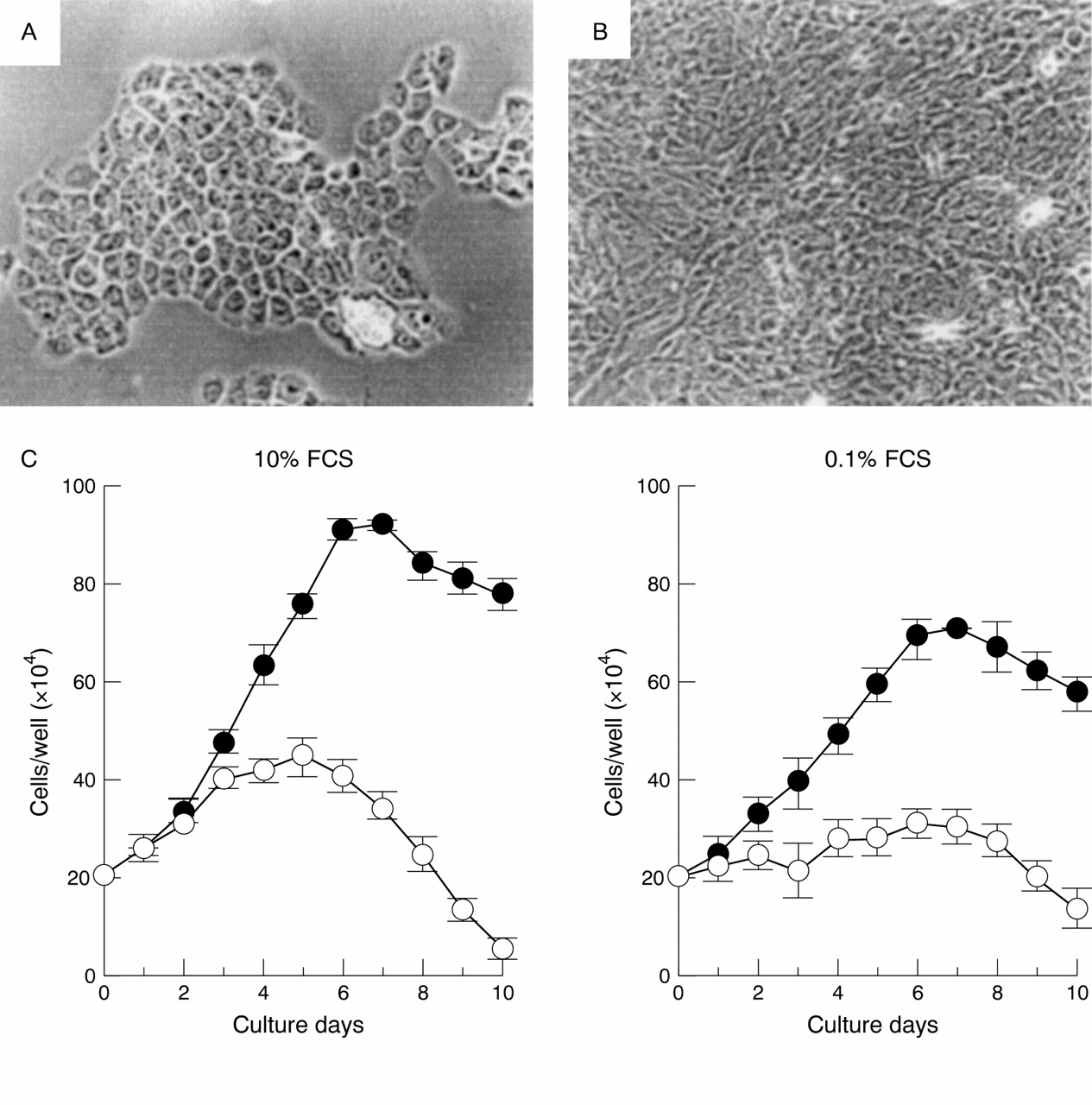
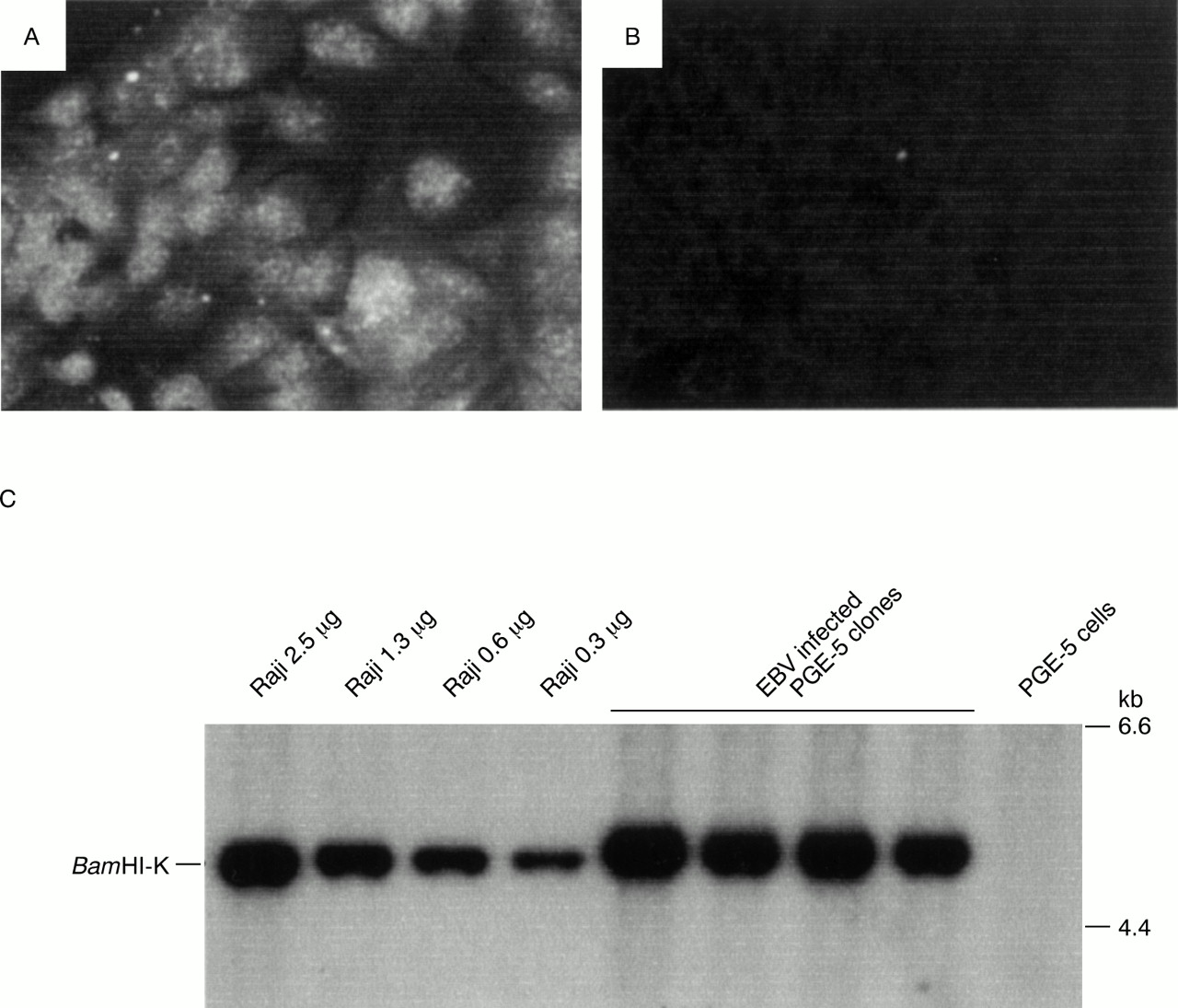
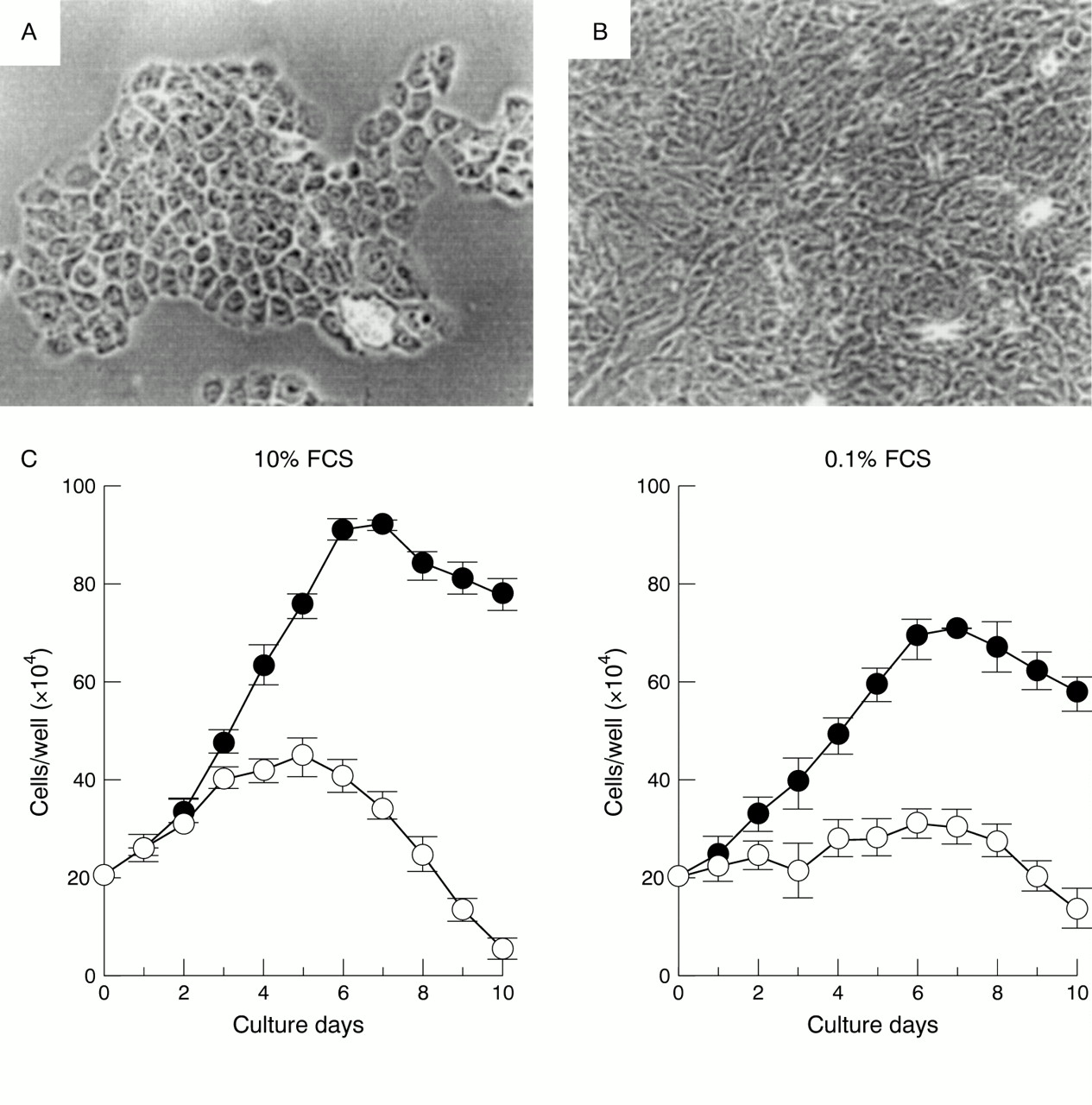
We attempted to infect gastric primary culture cells with EBV.65 Twenty one primary cell cultures from healthy gastric mucosa obtained by endoscopic biopsy were infected with EBV carrying a neomycin resistance gene (Neor), and infected cells were selected for using G418. As a result, EBNA expression (up to 0.2%) was observed in three of the cultures by three days after infection; in addition, from one culture we repeatedly separated cell clones that could be maintained for at least 300 generations. These cell clones were all 100% EBNA positive (fig 4) and expressed only the EBV genes EBNA1, EBER, BARF0, and LMP2A, similar to EBV positive gastric carcinomas. The EBV infected clones had higher proliferation rates and at least twice the cell saturation density in 10% serum compared with non-infected clones into which the Neor gene had been introduced as a control (fig 5), and the malignant phenotype was confirmed by colony formation in soft agar.

Detection of Epstein-Barr virus (EBV) in G418 resistant PGE-5 clones. (A) Immunofluorescent staining of EBV determined nuclear antigen (EBNA) in a G418 resistant PGE-5 clone with EBV seropositive human serum. Original magnification, ×400. (B) Immunofluorescent staining of the same clone with EBV seronegative human serum as a control. Original magnification, ×400. (C) Southern blot analysis of G418 resistant PGE-5 clones. All DNA samples were digested with BamHI and the blot was probed with a BamHI-K fragment of EBV DNA. Serially diluted Raji cell DNA served as positive controls. Each lane contained 5 μg of DNA. All G418 resistant PGE-5 clones were estimated to carry more than 25 copies of the EBV genome/cell. Reproduced from Nishikawa et al. 65
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Growth characteristics of neomycin resistance gene (Neor) transfected and Epstein-Barr virus (EBV) infected PGE-5 clones. (A) Neor transfected PGE-5 clone. Original magnification, ×100. (B) EBV infected PGE-5 clone. Original magnification, ×100. Both clones were detached by trypsinisation, seeded into separate wells of 12 well plates under the same culture conditions, and photographed near the plateau phase (five days after passage). Differences between the two cell types are easily recognisable. (C) Growth kinetics of EBV infected (closed circle) and Neor transfected (open circle) PGE-5 cells at normal (10%; left) and low (0.1%; right) fetal calf serum (FCS) concentrations. Each point represents the mean ±SE of the results for four clones. Reproduced from Nishikawa et al. 65
Normal gastric mucosal primary culture cells die after subculturing for a few generations, but one of the cultured cells from which the EBV infected clones were obtained was successively subcultured to a maximum of 60 generations without EBV infection. Therefore, the cells were not completely normal, but already had a functional abnormality in the p53 protein. These results suggest that EBV alone is not sufficient for oncogenesis, but can contribute to oncogenesis together with p53 or other gene abnormalities.
Differences in proliferation as a result of EBV infection were found not only in primary culture cells but also in some gastric carcinoma cell lines. The gastric carcinoma cell line NUGC3 showed the greatest difference, with a shorter cell doubling time, greater cell saturation density in 10% serum, and higher proliferation in soft agar. These results, together with the results for the primary culture cells, suggest that EBV expression in EBV positive gastric carcinomas contributes to cell malignancy.
Conclusions
It is clear that EBV is associated with some cases of gastric carcinoma. Thus far, many studies have focused on the mechanisms of immortalisation of primary B cells, and these studies revealed that LMP1 and EBNA2 are important for this process.60 On the other hand, in gastric carcinoma, in which a limited number of EBV genes are expressed, very little is known, largely because of the lack of a suitable in vitro system. We have developed a system of infecting epithelial cells with EBV, which simulates in vivo EBV expression.48,49 Comparison with uninfected cells has shown that the expression of a restricted set of EBV genes, which is similar to EBV positive gastric carcinoma, accelerates cellular proliferation and provokes malignancy.65 Thus, we have shown that EBV could contribute to the development of epithelial malignancies. A topic for future research will be to determine how and by what mechanism EBV causes these changes. We have developed a method of investigating this topic and are working towards an answer.
Note added in proof
The EBV BARF1 gene has been shown previously to act as an oncogene in several cell lines.66 Zur hausen et al have recently reported that the BARF1 gene is expressed in EBV carrying gastric adenocarcinomas,67 thus suggesting that the BARF1 gene might act as the viral oncogene in EBV carrying gastric carcinomas.